Se ha tenido la ocasión de comprobar de qué manera las formas tridimensionales de las proteínas les permiten desempeñar papeles estructurales y de transporte. Las enzimas son catalizadores biológicos selectivos de una eficiencia extraordinaria. Toda célula viva dispone de cientos de enzimas distintas que catalizan las reacciones esenciales para la vida. Aun los organismos vivos más simples contienen múltiples copias de cientos de enzimas diferentes. En los organismos multicelulares, el complemento de las enzimas varía de un tipo celular a otro.La mayor parte de las enzimas que se describen en este libro pertenecen a las más comunes que son virtualmente encontradas en todas las células. Estas enzimas catalizan las reacciones de las rutas metabólicas centrales, necesarias para mantener la vida.La mayor parte de las reacciones catalizadas por enzimas no procederían a velocidades apreciables bajo condiciones fisiológicas en ausencia de las enzimas. El papel principal de las enzimas es aumentar las velocidades de tales reacciones. En forma típica, las reacciones catalizadas por las enzimas son de 103 a 1020 veces más rápidas que las mismas sin catalizar.
Un catalizador es una sustancia que acelera la llegada a un equilibrio. Un catalizador puede cambiar en forma temporal durante la reacción, pero no cambia en el proceso general, porque se recicla para participar en varias reacciones. Los reactivos se unen a un catalizador y los productos se disocian de él. Un catalizador no cambia la posición del equilibrio de la reacción (es decir, no hace que una reacción no favorable sea favorable). Más bien reduce la cantidad de energía necesaria para que se efectúe la reacción.
Los catalizadores aceleran las reacciones tanto hacia adelante como hacia atrás al convertir un proceso de uno o dos pasos en varios pasos menores, cada uno con menor necesidad de energía que la reacción no catalizada. El capítulo siguiente describe los detalles de la forma en que las enzimas actúan como catalizadores.Las enzimas son muy específicas para los reactivos o sustratos sobre los que actúan, y varía el grado de especificidad hacia el sustrato.
• Pueden desempeñarse como catalizadores
• Catalizan reacciones muy específicas
• Pueden acoplar reacciones
• Su actividad puede ser regulada
El nombre enzima deriva de una palabra griega que significa “en la levadura”. Indica que dichos catalizadores están presentes en el interior de las células.
Los mecanicistas decían que las enzimas en las células de levadura eran las que catalizaban las reacciones de fermentación. Esta última conclusión fue respaldada por la observación de que los extractos de levadura, sin contenido de células, pueden catalizar la fermentación. A este hallazgo pronto siguió la identificación de las reacciones individuales y de las enzimas que las catalizan.
James B. Summer cristalizó, en 1926, la primera enzima ureasa y demostró que era una proteína. En la siguiente década se purificaron cinco enzimas más y se encontró que también eran proteínas: pepsina, tripsina, quimotripsina,
carboxipeptidasa y la enzima Old Yellow. Desde entonces se ha demostrado que casi todas las enzimas son proteínas, o proteínas más cofactores. Algunas moléculas de ARN también presentan actividad catalítica, pero usualmente no se les llama enzimas.






 Experimentos cinéticos revelan propiedades de las enzimas
Experimentos cinéticos revelan propiedades de las enzimas
El estudio de las propiedades de las enzimas se iniciara examinando las velocidades de reacciones catalizadas por las mismas. Tales estudios pertenecen a la categoría cinética enzimática
En los experimentos cinéticos se examina la relación entre la cantidad de producto (P) que se forma en una unidad de tiempo ( [P]/ t) y las condiciones experimentales bajo las que se efectúa la reacción. La base de la mayor parte de las mediciones cinéticas es la observación de la rapidez, o velocidad (v), de una reacción, la cual varía en forma directa con la concentración de cada reactante. Esta observación se expresa en una ecuación de velocidad. Por ejemplo, la ecuación de velocidad para la conversión no enzimática del sustrato (S) en el producto (P) en una reacción de isomerización.
La ecuación de velocidad refleja que la velocidad de una reacción depende de la concentración del sustrato ([S]). El símbolo k es la constante de velocidad e indica la velocidad o la eficiencia de una reacción. Cada reacción tiene una constante de velocidad diferente. Las unidades de la constante de velocidad para una reacción simple son s 1. Al avanzar una reacción, la cantidad de producto ([P]) aumenta y la cantidad de sustrato ([S]) disminuy.


Esta reacción se efectúa en dos pasos distintos: la formación del complejo enzimasustrato y la reacción química actual, acompañada por la disociación del producto. Cada paso transcurre con una velocidad característica. La velocidad total de una reacción enzimática depende de las concentraciones tanto del sustrato como del catalizador (la enzima). Cuando la cantidad de enzima es mucho menor que la cantidad de sustrato, la reacción depende de la cantidad de enzima.




Al suponer que la concentración de ES en estado estable es constante, entonces la velocidad de formación de producto depende de la velocidad de la reacción química y de la velocidad de disociación de P para abandonar la enzima. El paso limitante de la velocidad es hacia el lado derecho de la reacción y la velocidad depende de la constante de velocidad k2 y de la concentración de ES.
La derivación del estado estable resuelve la ecuación para [ES], usando términos que se pueden medir, como la constante de velocidad, la concentración total de la enzima ([E]total) y la concentración de sustrato ([S]). Se supone que [S] es mayor que [E]total,




Se ha estimado que un 20% de la población utiliza alguno de estos tipos de fármacos, hecho que ha dado un aumento del riesgo de padecer histaminosis alimentaria y por lo tanto desencadenar migraña, así como otros síntomas asociados a la intolerancia histamínica como molestias gastrointestinales, alteraciones dérmicas, dolores musculares, etc.

Analgésicos
Metamizol, Ácido acetil salicílico
Propiedades principales de las enzimas
• Pueden desempeñarse como catalizadores
• Catalizan reacciones muy específicas
• Pueden acoplar reacciones
• Su actividad puede ser regulada
-Casi todas son proteínas
-Son termosensibles
-Son hidrosolubles
-Pueden ser precipitadas
-Un 16% de su peso es nitrógeno
Los mecanicistas decían que las enzimas en las células de levadura eran las que catalizaban las reacciones de fermentación. Esta última conclusión fue respaldada por la observación de que los extractos de levadura, sin contenido de células, pueden catalizar la fermentación. A este hallazgo pronto siguió la identificación de las reacciones individuales y de las enzimas que las catalizan.
James B. Summer cristalizó, en 1926, la primera enzima ureasa y demostró que era una proteína. En la siguiente década se purificaron cinco enzimas más y se encontró que también eran proteínas: pepsina, tripsina, quimotripsina,
carboxipeptidasa y la enzima Old Yellow. Desde entonces se ha demostrado que casi todas las enzimas son proteínas, o proteínas más cofactores. Algunas moléculas de ARN también presentan actividad catalítica, pero usualmente no se les llama enzimas.
Las seis clases de enzimas
Los nombres en la mayor parte de las enzimas metabolicas se forman agregando el sufijo -asa- al nombre de sus sustratos, o a un termino descriptivo de la reaccion que catalizan. Ejemplo:
La ureasa tiene como sustrato la urea.
La alcohol deshidrogenasa cataliza la remocion de hidrogeno de los alcoholes.
Unas pocas enzimas, como la tripsina y la amilasa, se conocen por sus nombres historicos.
Muchas enzimas recién descubiertas reciben su nombre de acuerdo con sus genes o de alguna característica no descriptiva.
1. Las Oxidorreductasas:
Catalizan reacciones de oxido-reducción. Transferencia de hidrógeno o adición de oxígeno. (Lactato
deshidrogenasa)

EC 1.1.1.27
2. Las transferasas:
Catalizan las reacciones de transferencia de un grupo y pueden necesitar la presencia de coenzimas. El grupo transferido es diferente de hidrógeno.(Aminotransferasas, ej. Alanina transaminasa).
EC 2.6.1.2
3. Las hidrolasas:
Catalizan hidrólisis. Son una clase especial de transferasas donde el agua sirve como aceptor del grupo transferido. (Pirofosfatasa).

EC 3.6.1.1
4. Las liasas:
Catalizan la lisis de un sustrato, al generar un enlace doble, son reacciones de eliminación, no hidrolíticas y no oxidantes. (Piruvato descarboxilasa)

EC 4.1.1.1
5. Las isomerasas:
Catalizan cambios estructurales dentro de una misma molécula (reacciones de isomerización). Transferencia intra-molecular. (Alanina racemasa)

EC 5.1.1.1
6. Las ligasas:
Catalizan la ligadura o unión de dos sustratos. Este grupo de enzimas también reciben el nombre de sintetasas. Son dependientes de ATP. (Glutamina sintetasa).
EC 6.3.1.2
Distribución de todas las enzimas conocidas
1. Oxidorreductasa
2. Transferasas
3. Hidrolasas
4. Liasas
5. Isomerasas
6. Ligasas

El estudio de las propiedades de las enzimas se iniciara examinando las velocidades de reacciones catalizadas por las mismas. Tales estudios pertenecen a la categoría cinética enzimática
A. Cinética química
En los experimentos cinéticos se examina la relación entre la cantidad de producto (P) que se forma en una unidad de tiempo ( [P]/ t) y las condiciones experimentales bajo las que se efectúa la reacción. La base de la mayor parte de las mediciones cinéticas es la observación de la rapidez, o velocidad (v), de una reacción, la cual varía en forma directa con la concentración de cada reactante. Esta observación se expresa en una ecuación de velocidad. Por ejemplo, la ecuación de velocidad para la conversión no enzimática del sustrato (S) en el producto (P) en una reacción de isomerización.
La ecuación de velocidad refleja que la velocidad de una reacción depende de la concentración del sustrato ([S]). El símbolo k es la constante de velocidad e indica la velocidad o la eficiencia de una reacción. Cada reacción tiene una constante de velocidad diferente. Las unidades de la constante de velocidad para una reacción simple son s 1. Al avanzar una reacción, la cantidad de producto ([P]) aumenta y la cantidad de sustrato ([S]) disminuy.


Cinética enzimática
Uno de los primeros y grandes avances en bioquímica, fue el descubrimiento de que las enzimas se unen en forma transitoria a los sustratos, resultado de la investigación de la cinética enzimática. Emil Fischer, en 1894, propuso que una enzima presenta una plantilla rígida, o cerradura, y que el sustrato es la llave que le corresponde. Sólo los sustratos específicos se pueden ajustar a determinada enzima. Los primeros estudios de cinética enzimática confirmaron que una enzima (E) se une a un sustrato para formar un complejo enzima-sustrato (ES). Los complejos ES se forman cuando los ligandos se unen de manera no covalente a sus lugares adecuados en el sitio activo. El sustrato reacciona en forma transitoria con la proteína catalizadora (y con otros sustratos, en una reacción multisustratos) para formar el producto de la reacción. Imagínese ahora una reacción enzimática simple, la conversión de un sustrato en un producto catalizada por una enzima. Aunque la mayor parte de las reacciones enzimáticas tiene dos o más sustratos, se pueden describir los principios generales de la cinética enzimática suponiendo el caso sencillo en que hay un sustrato y un producto.Esta reacción se efectúa en dos pasos distintos: la formación del complejo enzimasustrato y la reacción química actual, acompañada por la disociación del producto. Cada paso transcurre con una velocidad característica. La velocidad total de una reacción enzimática depende de las concentraciones tanto del sustrato como del catalizador (la enzima). Cuando la cantidad de enzima es mucho menor que la cantidad de sustrato, la reacción depende de la cantidad de enzima.

Nota
-La velocidad total de una reacción enzimática depende de las
concentraciones tanto del sustrato como del catalizador (la enzima.)
-La velocidad de reacción esta
influenciada por la concentracion de E, pero no por la
concentración de otros reactantes, S.
Cinética química vs cinética enzimática
- Cinética enzimática depende de la [E],pero la E no es un S ni un P de la reacción.
- Las velocidades también son diferentes porque el sustrato debe unirse a la enzima para poder convertirse en producto.
Ecuación de Michaelis-Menten
Las reacciones catalizadas por enzimas, como cualquier reacción química, se pueden describir en forma matemática como ecuaciones de velocidad. En ellas, varias constantes indican la eficiencia y especificidad de una enzima y en consecuencia son útiles para comparar las actividades de varias enzimas o para evaluar la importancia fisiológica de una determinada enzima. Las primeras ecuaciones de velocidad fueron deducidas a principios de 1900 examinando los efectos de variaciones en la concentración de sustrato.
Los datos se pueden explicar con la reacción presentada en la ecuación. El primer paso es una interacción bimolecular entre la enzima y el sustrato para formar un complejo ES. A altas concentraciones de sustrato la velocidad inicial no cambia mucho cuando se agrega más S. Ello indica que la cantidad de enzima ha llegado a ser limitante de la velocidad de esta reacción. La concentración de la enzima es una parte importante de la reacción general, como es de esperarse para la formación de un complejo ES. A bajas concentraciones de sustrato, la velocidad inicial es muy sensible a cambios de concentración de sustrato. Bajo esas condiciones, la mayor parte de las moléculas de enzima todavía no se han unido al sustrato y la formación del complejo ES depende de la concentración de sustrato.
Se observó que la forma de la curva describía una HIPÉRBOLA


La enzima está a la mitad de la saturación cuando S=Km
Esta es llamada la ecuación de Michaelis-Menten, por Leonor Michaelis y Maud Menten. La ecuación de Michaelis-Menten describe la relación entre la velocidad inicial de una reacción y la concentración del sustrato. En la sección que sigue se deducirá la ecuación de Michaelis-Menten por un método cinético para después examinar el significado de las diversas constantes.
A. Deducción de la ecuación de Michaelis-Menten
Una deducción común de la ecuación de Michaelis-Menten, debida a George E. Briggs y J. B. S. Haldane, es llamada la derivación del estado estable. Esta deducción postula que hay un intervalo de tiempo (llamado estado estable, o estado estacionario) durante el cual se forma el complejo ES a la misma velocidad con la que se descompone, de modo que la concentración de ES es constante. La velocidad inicial se usa en la derivación del estado estable porque se asume que la concentración de producto ([P]) es insignificante. Tal estado estable es una condición común para las reacciones metabólicas en las células.Al suponer que la concentración de ES en estado estable es constante, entonces la velocidad de formación de producto depende de la velocidad de la reacción química y de la velocidad de disociación de P para abandonar la enzima. El paso limitante de la velocidad es hacia el lado derecho de la reacción y la velocidad depende de la constante de velocidad k2 y de la concentración de ES.
La derivación del estado estable resuelve la ecuación para [ES], usando términos que se pueden medir, como la constante de velocidad, la concentración total de la enzima ([E]total) y la concentración de sustrato ([S]). Se supone que [S] es mayor que [E]total,

B.Constante catalítica kcat
Cuando la concentración de sustrato es alta, la velocidad total de la reacción es Vmáx y está determinada por la concentración de la enzima. La constante de velocidad observada bajo estas condiciones se llama constante catalítica, Kcat, y se define como sigue:

Donde kcat representa la cantidad de moles de sustrato convertidos en producto, por segundo y por mol de enzima (o por mol de sitio activo, para una enzima con multisubunidades) bajo condiciones de saturación. En otras palabras, kcat indica la cantidad máxima de moléculas de sustrato convertidas en producto cada segundo por cada sitio activo. A eso se le llama con frecuencia número de recambio. La constante catalítica mide la rapidez con que determinada enzima puede catalizar una reacción específica; es una forma muy útil para describir la eficacia de una enzima. La unidad de kcat es s 1. El recíproco de kcat es el tiempo necesario para que haya un evento catalítico. Nótese que para obtener kcat se debe conocer la concentración de la enzima.
C. Significados de Km
La constante de Michaelis tiene varios significados. Define a Km como la relación de las constantes de velocidad combinadas para la descomposición de ES dividida entre la constante para su formación. Si la constante de velocidad para la formación de producto (k2) es mucho menor que k1 o que k 1, como es el caso frecuente, se puede despreciar k2 y Km equivale a k 1/k1. En este caso, Km es igual a la constante de equilibrio de disociación de ES para dar E S. Así, Km es una medida de la afinidad de E hacia S. Mientras menor sea el valor de Km, el sustrato está más fuertemente unido. Km también es uno de los parámetros que determina la forma de la curva de y0 en función de [S] de la figura 5.4b. Es la concentración del sustrato cuando la velocidad inicial es la mitad del valor de Vmáx. Este significado es consecuencia directa de la ecuación general de una hipérbola rectangular.
Medición de Km y Vmáx
Los parámetros cinéticos de una reacción enzimática pueden producir información valiosa acerca de la especificidad y el mecanismo de la reacción. Los parámetros clave son Km y Vmáx ya que kcat se puede calcular si se conoce Vmáx.
Los datos de Km y Vmáx para una reacción catalizada por enzima se pueden determinar de diversas maneras. Es posible obtener ambos valores por análisis de las velocidades iniciales en una serie de concentraciones de sustrato y una concentración fija de enzima. Para obtener valores fiables de las constantes cinéticas, los puntos de [S] se deben extender por abajo y por arriba de Km para producir una hipérbola. La ecuación de Michaelis-Menten se puede reacomodar para obtener valores de Vmáx y Km a partir de líneas rectas en gráficas. La transformación de uso más frecuente es la gráfica de doble recíproco, o de Lineweaver-Burk, en la que se grafican los valores de 1/y0 contra los de 1/[S]. El valor absoluto de 1/Km se obtiene con la intersección de la recta con el eje x (es decir, la abscisa al origen), y el valor de 1/Vmáx se obtiene con la ordenada al origen. Aunque las gráficas de doble recíproco no son los métodos más exactos para determinar las constantes cinéticas, se comprenden con facilidad y permiten contar con pautas reconocibles para estudiar la inhibición enzimática, un aspecto de extrema importancia en la enzimología que en breve se examinará. Se pueden obtener valores de kcat con mediciones de Vmáx sólo cuando se conoce la concentración absoluta de la enzima. Se pueden determinar los valores de Km aun con enzimas que no hayan sido purificadas siempre y cuando sea una sola la enzima en la preparación impura la que pueda catalizar la reacción observada.
Cinética de las reacciones con sustratos múltiples
Las mediciones cinéticas de reacciones con sustratos múltiples (o reacciones de multisustrato) son algo más complicadas que para cinéticas enzimáticas sencillas con un solo sustrato. Sin embargo, para muchos fines, como por ejemplo el diseño de una cuantificación enzimática, basta sólo con determinar la Km para cada sustrato en presencia de cantidades saturantes de cada uno de los demás sustratos, como fue descrito para las reacciones químicas.
A. Reacciones Consecutivas
En las Rx consecutivas, todos los sustratos se enlazan antes de liberar un producto. La unión de los sustratos puede ser ordenada o aleatoria.

b) Reacción de ping-pong de bisustrato
En las reacciones ping-pong, un sustrato es enlazado y un producto es liberado dejando una enzima sustituida. Entonces se enlaza un segundo sustrato y se libera un segundo producto, con lo cual la enzima regresa a su forma original.

¿Has oído sobre inhibidores enzimáticos? ¿Sabes lo que son? ¿Como Actúan?
¿Te gustaría aprender sobre ellos?
Existes Dos grandes grupos
Click Click
Mira este vídeo acerca de Inhibidores Enzimáticos
Medicamento inhibidor de enzima
Farmacos Bloqueadores de DAO
Existe un conjunto de fármacos implicados en el déficit o en la baja actividad de la enzima Diamino Oxidasa. Bloquean o inhiben enzimas implicadas en la metabolización de la histamina, sobretodo de la DAO, o liberan histamina endógena. Este riesgo es de gran importancia, ya que se han publicado alrededor de 90 fármacos involucrados, algunos de los cuales de uso muy común.Se ha estimado que un 20% de la población utiliza alguno de estos tipos de fármacos, hecho que ha dado un aumento del riesgo de padecer histaminosis alimentaria y por lo tanto desencadenar migraña, así como otros síntomas asociados a la intolerancia histamínica como molestias gastrointestinales, alteraciones dérmicas, dolores musculares, etc.
Analgésicos
Metamizol, Ácido acetil salicílico

Especialidad Analgesia Sitio y mecanismo de acciónEl metamizol (dipirona) es un analgésico-antipirético del grupo de las pirazolonas; se considera como dervicado soluble de la aminopirina y comparte con ésta los riesgos de producir agranulocitosis. También tiene propiedades antiinflamatorias y espasmolíticas. Inhibe la acción de la ciclooxigenasa, y en consecuencia la síntesis de prostaglandinas, acción que parece explicar sus propiedades analgésicas y antipiréticas. Es posible que su efecto analgésico también dependa de una acción central. Por otro lado, relaja y reduce la actividad del músculo liso gastrointestinal y uterino. Cinética Se absorbe bien después de administración oral y su concentración plasmática alcanza un nivel máximo entre los 30 y 120 min. Puede ser administrado por vía intramuscular o intravenosa. Tiene una vida media biológica de 8 a 10 h. Su unión a proteínas plasmáticas es discreta, la metilaminoantipirina se metaboliza en el hígado a aminoantipirina y es eliminada por riñón en 90%. Indicaciones Adultos: Dolor moderado a intenso cuando no puede ser tratado con otros analgésicos menos tóxicos. Niños: No se recomienda su empleo en menores de 12 años. Dosis Oral. 1 a 2 g cada 8 h. Intramuscular (intraglútea profunda): 1 g cada 6 a 8 h. Intravenosa. 1 a 2 g cada 12 h. Preparar como indica el fabricante y administrar muy lentamente (no más de 1 ml/min). El paciente deberá estar en decúbito, y de preferencia con registro de presión arterial, frecuencia cardiaca y respiración. |
Ritonavir
El ritonavir es un antirretroviral del grupo de los inhibidores de la proteasa y se utiliza en la terapia recombinante antirretroviral activa de adultos y pacientes pediátricos con infección por VIH-1 y enfermos de sida. Este medicamento fue aprobado por la FDA el 1 de marzo de 1996, siendo el séptimo de su tipo que se aprobó.Mecanismo de acción
Este medicamento se utiliza en la terapia recombinante para la reducción de la carga viral del VIH inhibiendo a la proteasa, aunque recientemente se utiliza también como inhibidor del citocromo o CYP3A, que metaboliza otros medicamentos antirretrovirales como el lopinavir, con lo que aumenta la concentración del medicamento en la sangre y su potencialidad en el organismo.
Vía de administración y efectos adversos
Su administración se hace por vía oral, tomado junto con los alimentos. Su efecto adverso más importante es la hiperglucemia, debido a que inhibe los mediadores de la insulina, por lo que puede llegar a ocasionar problemas a los enfermos de diabetes mellitus de tipo 1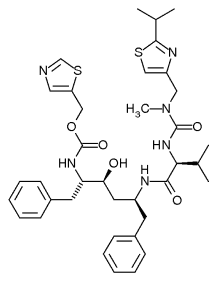
IECA (INHIBIDORES DE LA ENZIMA CONVERTIDORA DE LA ANGIOTENSINA)
¿Qué hacen?Los inhibidores de la enzima convertidora de la angiotensina (IECA) bloquean los efectos de una hormonaproducida naturalmente por los riñones denominadaangiotensina II. Al bloquear el efecto de la angiotensina II, los IECA relajan los vasos sanguíneos, lo que disminuye la presión arterial. Esto significa que su corazón no tiene que trabajar tanto para llevar la sangre al organismo.
Reacciones adversas
Como los IECA disminuyen la presión arterial, a veces puede sentirse mareado. Se vigilará con regularidad su presión arterial. Los IECA también modifican ligeramente la función renal o aumentan las concentraciones de potasio - su médico lo comprobará con regularidad mediante análisis de sangre.
Algunas personas también tienen una tos seca. Si esto le ocurre a usted, el médico le reducirá la dosis o le cambiará la medicación. Con menos frecuencia, las personas presentan alteraciones del gusto o erupciones cutáneas. Muy rara vez, las personas que toman IECA sufren hinchazón de los labios o de la garganta. En caso de que esto suceda, avise inmediatamente a su médico.
Consejos básicos
Su médico puede aconsejarle que tome la primera dosis de su IECA antes de acostarse, para así evitar un posible mareo. El mareo es frecuente con cualquier medicamento que disminuya la presión arterial, pero su corazón puede funcionar mejor con estas presiones más bajas.
Benazeprilo


DESCRIPCION
El benazepril es un inhibidor de la enzima de conversión de la angiotensina, activo por vía oral, usado en el tratamiento de la hipertensión. El benazepril es un profármaco que es hidrolizado por las esterasas del plasma al benazeprilato, cuya larga duración lo hace adecuado para un tratamiento a base de dosis únicas al día. Farmacodinámicamente, el benazepril se asemeja al fosinopril, quinapril, y ramipril. Ni el benazepril ni el benazeprilato contienen grupos sulfihidrilo, lo que explica el bajo nivel de reacciones adversas de este fármaco.
Mecanismo de acción: el benazepril compete con la enzima de conversión de la angiotensina (ECA) frente al sustrato de angiotensina I, bloqueando su conversión a angiotensina II. La angiotensina II es un potente vasoconstrictor y un factor de retroalimentación negativo de la actividad de la renina. Al reducir los niveles plasmáticos de angiotensina II, el benazepril reduce la presión arterial aumentando la actividad de la renina. Además, en respuesta a la caída de la presión arterial los mecanismos baroreflejos son estimulados. Los inhibidores de la ECA reducen pues el tono vascular al reducir localmente la actividad vasoconstrictora de la angiotensina II. La disminución de los niveles plasmáticos de angiotensina II también reduce la secreción de aldosterona, disminuyendo la retención de agua y de sodio. El benazepril produce una vasodilatación arterial reduciendo las resistencias periféricas. En los pacientes hipertensos, la presión arterial es reducida sin que se produzcan cambios en el volumen-latido, frecuencia cardíaca o gasto cardíaco. Después de la administación del benazepril disminuyen tanto la presión arterial supina como de pie, y aunque la hipotensión sintomática rara vez se produce puede ocurrir en pacientes hiponatrémicos o hipovolémicos. El benazepril, al igual que los demás inhibidores de la ECA no afectan la resistencia insulínica, no alteran el perfil lipoproteíco ni ocasionan disfunción sexual.
Farmacocinética: el benazepril se administra por vía oral, siendo rápidamente absorbido por el tracto gastrointestinal. Los alimentos no afectan la biodisponibilidad de este fármaco. El benazepril es un profármaco del benazeprilato cuyas concentraciones plasmáticas máximas son obtenidas a las 1-2 horas después de la administración del fármaco en ayunas y a las 2-4 horas si se ingiere con alimentos. Para generar el metabolito activo se requiere una metabolización hepática. Los efectos cardiovasculares comienzan a aparecer al cabo de una hora, siendo máxima la reducción de la presión arterial al cabo de 2 a 4 horas. Los efectos hipotensores duran aproximadamente 24 horas, permitiendo un régimen de tratamiento de una dosis única al día. El fármaco se une a las proteínas del plasma en un 95% y muestra un volumen de distribución reducido. El benazepril es convertido en su totalidad al metabolito activo, siendo este último eliminado por vía real en su mayoría y tan solo en un 10% por vía biliar. La semivida efectiva del benazeprilato es de 10-11 horas. La farmacocinética del benazepril es afectada por el estado de la función renal, pero no por la edad o por la disfunción hepática. La biodisponibilidad del benazepril en los pacientes con disfunción renal moderada (aclaramiento de creatinina > 30 ml/min) es similar a la de los pacientes normales, aumentando los niveles plasmáticos y la duración de la fase a de eliminación en pacientes con insuficiencia renal grave
Enzimas alostéricas
Son invariablemente proteínas con múltiples subunidades, con múltiples lugares activos. Son enzimas cuyas propiedades son afectadas por cambio en la estructura.
Cinética sigmoide, indica cooperatividad entre las subunidades. La unión de un sustrato a uno de los sitios, afecta el enlace en los otros sitios.
Son reguladas por la unión de:
• Moduladores positivos
• Moduladores negativos que pueden ser:
-Homotrópicos o homoalosterismo
-Heterotrópicos o heteroalosterismo

Propiedades generales de las enzimas alostéricas
1. La actividad de las enzimas alostéricas cambian debido a inhibidores y activadores metabólicos. Con frecuencia los moduladores alostéricos no se parecen a los sustratos o a los productos de la enzima.
2. Los moduladores alostéricos se enlazan en forma No covalente a las enzimas que regulan. Los moduladores mismos no son alterados químicamente por la enzima.
3. Con pocas excepciones, las enzimas reguladoras son proteínas de subunidades múltiples. Las cadenas polipeptídicas individuales de una enzima reguladora pueden ser idénticas o diferentes.
4. Toda enzima sujeta a regulación alostérica posee cuando menos un sustrato para el cual la curva de Vo en función de [S] es sigmoidea en lugar de hiperbólica.
Cooperatividad
Las proteinas con efectos cooperativos tienen curvas de saturacion deferrentes a la hiperbola rectangular (Michaelis-Menten)
Modelo de Cooperatividad concertado
- Referido Tambien como el modelo de la conservacion de la simetria propuesto por Monod, Wymann y Changeux
- Postula que las subunidades estan conectadas que el cambio conformacional en una subunidad se confiere a todas subunidades
- Todas las subunidades estan en la misma conformacion
- La union del ligando (sustrato o modulador) favorece el equilibrio de uno de los estados conformacionales T o R
- El equilibrio entre los estados R y T es FAvorecido Por la union del Ligando

Modelo de Cooperatividad secuencial
- El modelo secuencial de la regulación alostérica sostiene que las subunidades están conectadas en forma que el cambio conformacional en una subunidad no induce en cambio similar en las otras.
- Todas las subunidades de la enzima no están necesariamente en la misma conformación
- La union del sustato a la enzima es via de adecuacion inducida, que indica que cuando una molecula de sustrato colisiona con el sitio activo, la conformacion del sitio activo cambia para unir al sustrato
- Esta union propada el cambio en las subunidades adyacentes que aumentan la afinidad por el sustrato, no necesariamente a todas

Actividades
Aplique la ecuación de Michaelis-Menten para demostrar lo siguiente:
a) Vo se vuelve independiente de [S] cuando [S] >> Km.
b) La reacción es de primer orden con respecto a S cuando
[S] << Km.
c) [S] = Km cuando Vo es la mitad de Vmáx.
Trace una gráfica de contra [S] para una reacción enzimática típica
a) en ausencia de inhibidor
b) en presencia de un inhibidor competitivo
c) en presencia de un inhibidor no competitivo.
No hay comentarios.:
Publicar un comentario